
北方农牧交错区干旱特征变化及其对植被总初级生产力的影响
|
周温存(1997— ),女,江西九江人,硕士生,研究方向为遥感与地理信息系统应用。E-mail: 2020110334@ecut.edu.cn |
收稿日期: 2022-06-08
修回日期: 2022-08-19
网络出版日期: 2023-04-19
基金资助
国家自然科学基金项目(41971218)
中国科学院A类战略性先导科技专项(XDA23070302)
Spatio-temporal Changes of Drought Features and Their Impacts on the Gross Primary Production in Farming-Pastoral Ecotone of Northern China
Received date: 2022-06-08
Revised date: 2022-08-19
Online published: 2023-04-19
Supported by
National Natural Science Foundation of China(41971218)
The Strategic Priority Research Program (Class A) of the Chinese Academy of Sciences(XDA23070302)
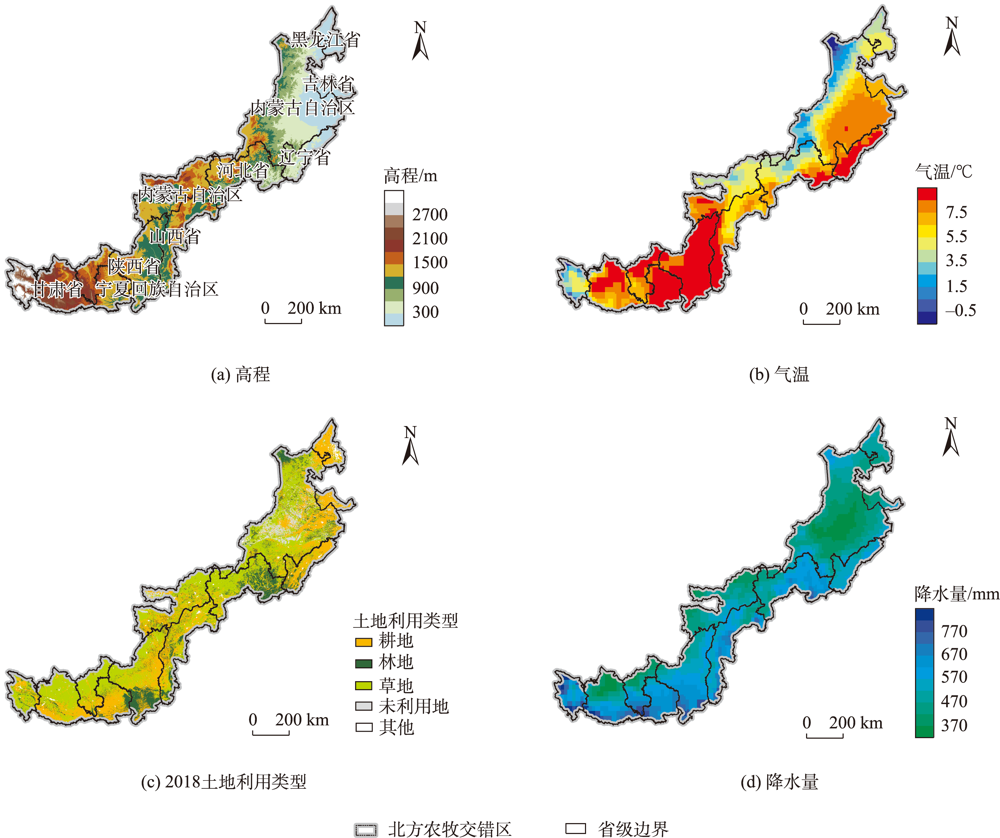
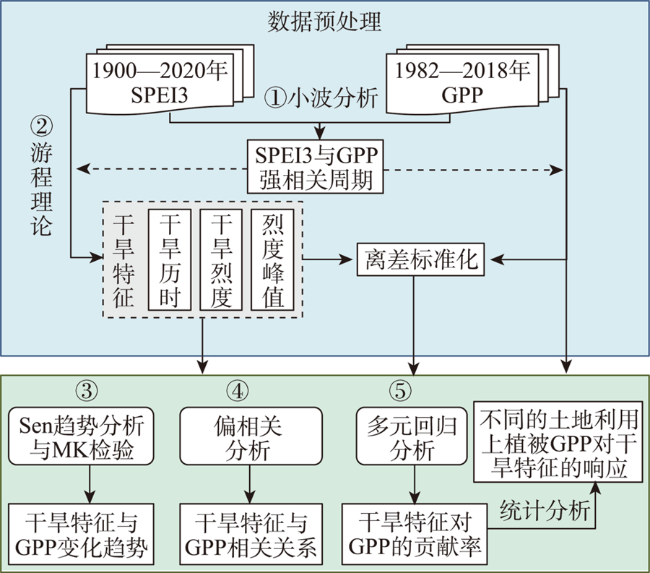
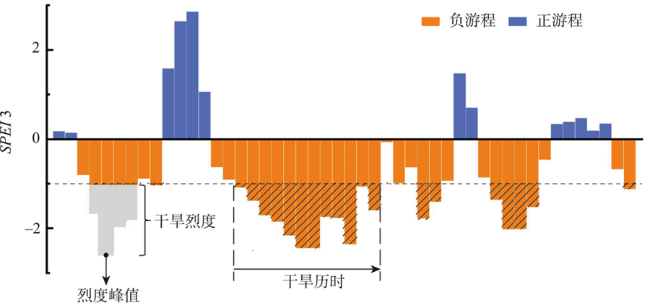
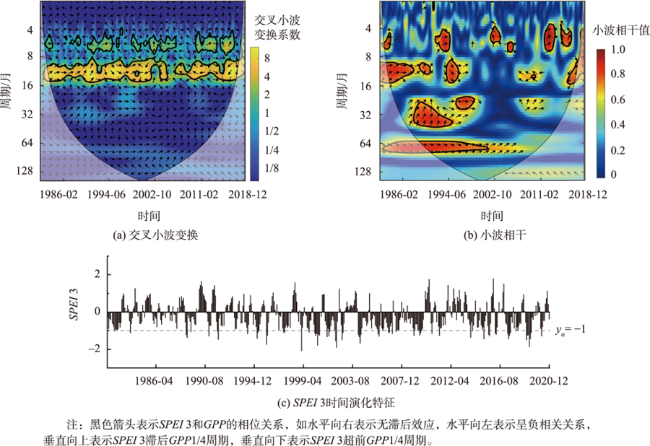
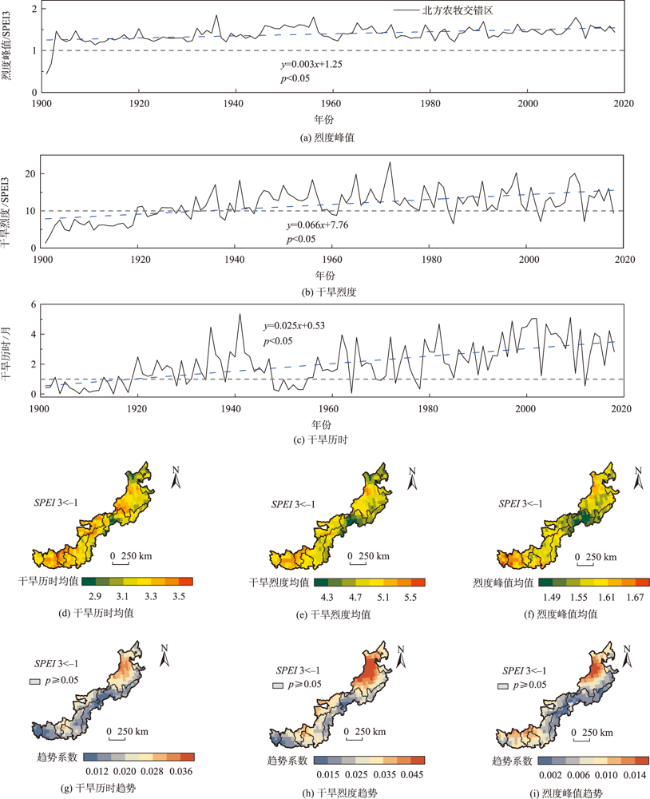
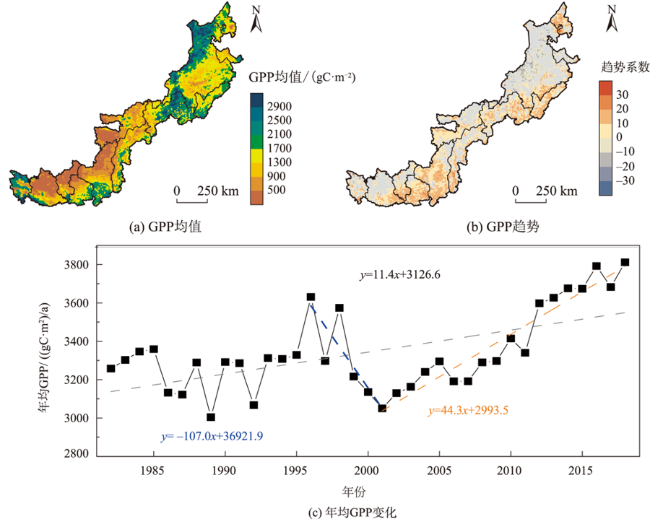
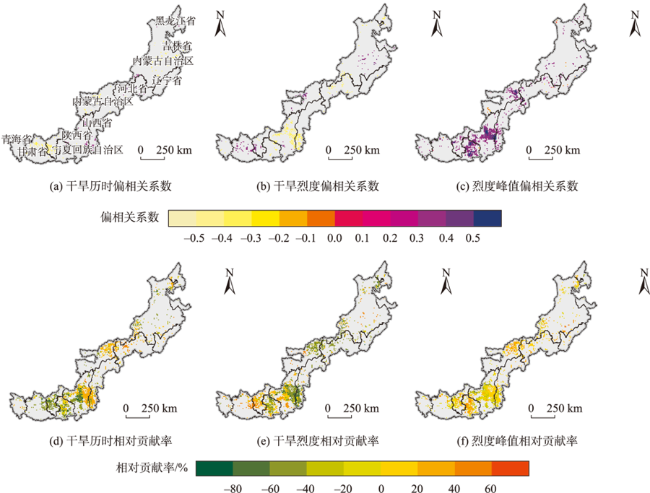
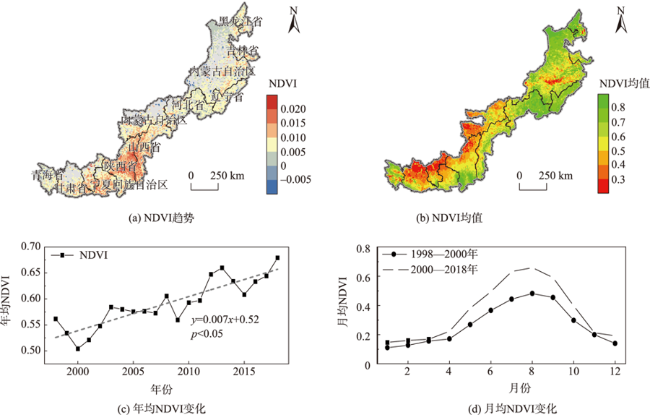
北方农牧交错区地处半湿润/半干旱生态脆弱过渡带,干旱是影响该区植被生产力的关键因素之一。探究干旱对植被总初级生产力的影响,对深刻理解气候变化下生态系统生产力变化响应特征及优化区域碳水循环具有重要意义。为了更好地了解水分限制区不同干旱特征对GPP影响,本研究以北方农牧交错区为例,基于长时间序列的标准化降水蒸散发指数(SPEI3,1900—2020年)和植被总初级生产力(GPP,1982—2018年)等数据,首先采用小波分析明确SPEI3与GPP强相关周期,在此基础上利用游程理论识别干旱特征,进而分析了北方农牧交错区干旱特征与GPP的变化趋势,最后厘定了不同干旱特征对GPP的影响。结果表明:① 1982—2018年北方农牧交错区SPEI3与GPP在半年周期和年周期存在显著相关关系,滞后效应随时间变化而变化;年际分析能够减弱滞后效应对SPEI3与GPP相关性的影响;② 1900—2020年北方农牧交错区干旱历时、干旱烈度和烈度峰值均呈现显著增加趋势,干旱烈度随着干旱历时和烈度峰值的增加而加剧,干旱特征高值区往往具有更强的增加趋势;③ 1982—2018年北方农牧交错区GPP总体呈现增加趋势,GPP高值区表现出更强的增加趋势;④ 不同干旱特征对GPP变化的影响不同,贡献率绝对值表现为干旱烈度>干旱历时>烈度峰值;整体来看,干旱特征共同解释了GPP变化面积的18.1%,干旱历时和干旱烈度显著抑制GPP增加;⑤ 不同的土地利用上植被GPP变化对干旱特征响应不同,乔木和灌木GPP下降主要来自干旱历时的负贡献,草地和耕地GPP下降则由干旱烈度的负贡献主导。
周温存 , 刘正佳 , 王坤 , 邹时林 , 钟会民 , 陈芳鑫 . 北方农牧交错区干旱特征变化及其对植被总初级生产力的影响[J]. 地球信息科学学报, 2023 , 25(2) : 421 -437 . DOI: 10.12082/dqxxkx.2023.220390
Climate change has been projected to increase the drought duration and intensity in the future, with greater impact on the Gross Primary Production (GPP) in areas. However, despite our increasing understanding of the drought impacts on GPP in recent decades, there is still little understanding of how different drought features (i.e., duration, intensity, peak) GPP. A better understanding of the impact of different drought features on GPP under climate change can help optimize the regional carbon water cycle. In this study, we took the Farming-Pastoral Ecotone of Northern (FPEN) in China as an example and used long-term time series Standardized Precipitation Evapotranspiration Index (SPEI3) during 1900—2020 and GPP during 1982—2018. Firstly, the cross wavelet transform and wavelet coherence analysis was employed to reveal the correlation relationship between SPEI3 and GPP at different temporal scales. Then, the drought features were identified by runs theory, and the trend of drought features and GPP were analyzed using the Theil-Sen Median trend analysis with the Mann-Kandall test in FPEN. Finally, the effects and relative contributions of different drought features on GPP were determined by partial correlation analysis and multiple linear regression analysis. The results show that: (1) There was a strong correlation between SPEI3 and GPP at the scales of half year (6±1 month) and 1 year (12±2 month) in FPEN during 1982—2018, and the 1-year scale had a stronger relationship. Meanwhile the lag effect of SPEI3 and GPP changed with time. Therefore, we clarified that the interannual analysis was able to reduce the effect of lag effect on the correlation between SPEI3 and GPP; (2) The drought duration, intensity, and peak all showed a significantly increasing trend in FPEN during 1900—2020. Drought intensity increased with increasing drought duration and peak. The drought features with higher values often had a stronger increasing trend; (3) GPP showed a trend of “increase-decrease-increase”, with a significant increasing trend during 1982—2018, and the high values of GPP showed a stronger increasing trend; (4) Different drought features have different effects on GPP changes. The absolute contribution rate of drought features was in the order of intensity > duration > peak. Overall, drought features together explained 18.1% of the GPP change. Drought duration and intensity significantly inhibited GPP growth, with the relative contribution rate of -0.05 and -0.35, respectively; (5) GPP of different land covers responded differently to drought features. The drought duration and intensity has negative contribution to the GPP changes in forests/shrubs and grasslands/croplands, respectively.
表1 本文研究使用数据说明Tab. 1 Data description |
| 数据类型 | 时间 | 空间分辨率 | 时间分辨率 | 来源 |
|---|---|---|---|---|
| 总初级生产力 | 1982—2018年 | 0.5° | 逐月 | https://doi.org/10.1016/j.scitotenv.2020.142569 |
| 标准化降水蒸散发指数 | 1900—2020年 | 0.5° | 逐月 | https://spei.csic.es/spei_database |
| 高程 | 2000年 | 30″ | - | https://earthdata.nasa.gov/ |
| 植被归一化指数 | 1998—2018年 | 1 km | 逐月 | https://www.resdc.cn/ |
| 温度 | 1992—2020年 | 0.25° | 逐月 | https://cds.climate.copernicus.eu |
| 降水 | 1992—2020年 | 0.25° | 逐月 | https://cds.climate.copernicus.eu |
| 土地利用数据 | 2018年 | 300 m | 逐年 | https://cds.climate.copernicus.eu |
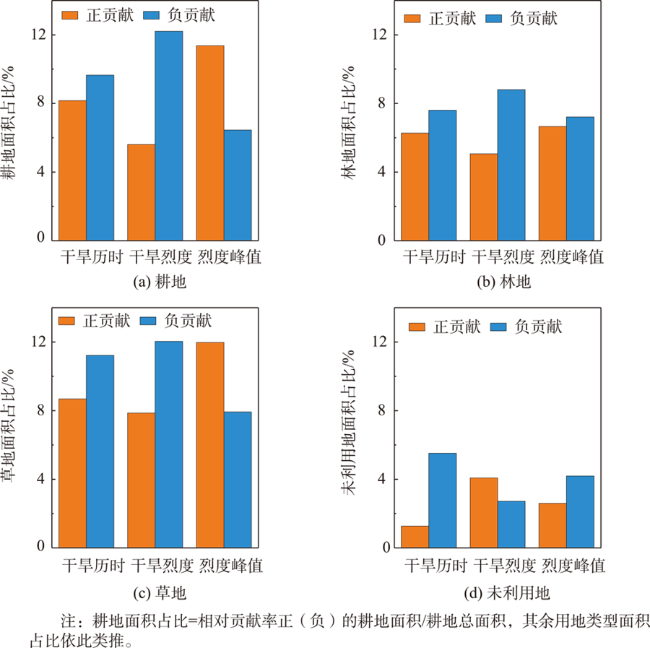
图7 1982—2018年北方农牧交错区干旱特征对GPP变化的影响与贡献率Fig. 7 Effects and Contributions of drought features on GPP changes in the farming-pastoral ecotone of northern China from 1982 to 2018 |
表2 1982—2018年北方农牧交错区干旱特征与GPP的相关关系以及干旱特征对GPP变化的相对贡献率Tab. 2 The correlation between drought features and GPP and relation contributions of drought features on GPP changes in the farming-pastoral ecotone of northern China from 1982 to 2018 |
| 变量 | 偏相关分析模型 | 多元回归模型 | |||||
|---|---|---|---|---|---|---|---|
| 偏相关系数 | 正相关面积/km2 | 负相关面积/km2 | 相对贡献率/% | 正贡献面积/km2 | 负贡献面积/km2 | ||
| 干旱历时 | -0.20* | 9477 | 27 621 | -13.95* | 58 239 | 73 629 | |
| 干旱烈度 | -0.17* | 12 231 | 29 808 | -20.44* | 48 762 | 83 106 | |
| 烈度峰值 | 0.38* | 68 850 | 3078 | 4.16* | 77 922 | 53 946 | |
注:*表示显著性检验p<0.05。 |
表3 2018年北方农牧交错区不同土地利用类型上干旱特征对GPP的相对贡献率Tab. 3 Relation contributions of drought features to GPP on different land use types in the farming-pastoral ecotone of northern China in 2018 |
| 植被类型 | 相对贡献率/% | |||
|---|---|---|---|---|
| 干旱历时 | 干旱烈度 | 烈度峰值 | ||
| 耕地 | -12.70* | -15.19* | 10.94* | |
| 林地 | 乔木 | -39.06* | -6.22* | 5.43* |
| 灌木 | -17.11* | -12.67* | 9.07* | |
| 草地 | -12.59* | -16.71* | 12.35* | |
注:*表示显著性检验p<0.05。 |
| [1] |
孙颖. 人类活动对气候系统的影响——解读IPCC第六次评估报告第一工作组报告第三章[J]. 大气科学学报, 2021, 44(5):654-657.
[
|
| [2] |
周波涛. 全球气候变暖:浅谈从AR5到AR6的认知进展[J]. 大气科学学报, 2021, 44(5):667-671.
[
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
张世喆, 朱秀芳, 刘婷婷, 等. 气候变化下中国不同植被区总初级生产力对干旱的响应[J]. 生态学报, 2022, 42(8):3429-3440.
[
|
| [20] |
|
| [21] |
靖娟利, 和彩霞, 王永锋, 等. 西南地区1902—2018年干旱时空演变特征分析[J]. 水土保持研究, 2022, 29(3):220-227.
[
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
原媛, 母艳梅, 邓钰洁, 等. 植被覆盖度和物候变化对典型黑沙蒿灌丛生态系统总初级生产力的影响[J]. 植物生态学报, 2022, 46(2):162-175.
[
|
| [27] |
王大为, 周伟, 韩涛, 等. 2000—2019年石羊河流域植被总初级生产力变化及其气候特征[J/OL]. 生态学杂志.https://kns.cnki.net/kcms/detail/21.1148.Q.20220617.1438.010.html.
[
|
| [28] |
|
| [29] |
王永财, 孙艳玲, 王中良. 1998—2011年海河流域植被覆盖变化及气候因子驱动分析[J]. 资源科学, 2014, 36(3):594-602.
[
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
董伯纲, 于洋. 近60年山西省极端降水时空变化特征[J]. 水土保持学报, 2022, 36(1):135-41.
[
|
| [46] |
|
| [47] |
冯瑞瑞, 荣艳淑, 吴福婷. 基于Copula函数的宜昌水文干旱特征分析[J]. 水文, 2020, 40(2):23-30,71.
[
|
| [48] |
杨少康, 刘冀, 魏榕, 等. 长江上游流域生长季气象干旱分异特征[J]. 水土保持研究, 2022, 29(2):184-191.
[
|
| [49] |
钟洋洋, 钱诚. 中国地区春霜冻的变化趋势和未来情景预估[J]. 气候与环境研究, 2022, 27(1):50-62.
[
|
| [50] |
张彦, 程锐, 邹磊, 等. 气候变化和人类活动对灌区地下水埋深的影响[J]. 灌溉排水学报, 2022, 41(2):91-100.
[
|
| [51] |
李肖,
[
|
| [52] |
尤南山, 董金玮, 肖桐, 等. 退耕还林还草工程对黄土高原植被总初级生产力的影响[J]. 地理科学, 2020, 40(2):315-323.
[
|
| [53] |
|
| [54] |
|
| [55] |
王丽霞, 史园莉, 张宏伟, 等. 2000—2020年北方农牧交错区植被生态功能变化及驱动因子分析[J]. 生态环境学报, 2021, 30(10):1990-1998.
[
|
| [56] |
杜金燊, 于德永. 气候变化和人类活动对中国北方农牧交错区草地净初级生产力的影响[J]. 北京师范大学学报(自然科学版), 2018, 54(3):365-372.
[
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
罗丹丹, 王传宽, 金鹰. 植物水分调节对策:等水与非等水行为[J]. 植物生态学报, 2017, 41(9):1020-1032.
[
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}