
Journal of Geo-information Science >
An Analysis of the Spatial Variations and Seasonal Changes of Methane Emission from Cattle in China
Received date: 2014-04-09
Request revised date: 2015-05-12
Online published: 2015-09-07
Copyright
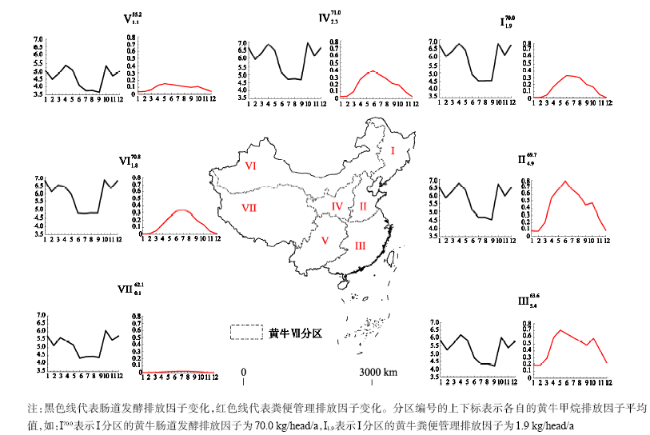
Ruminant livestock accounts for the major proportion of methane emissions within the agricultural sector. In China, cattle dominates the livestock due to its huge population and large size in comparison to sheep. The latest studies have paid little attention to the spatial variations and seasonal changes of the livestock methane emission factors, though a lot of direct measurements and modeling estiamtions have been made to improve the quality of the national inventories. In this study, we analyzed the spatial variarions and seasonal changes of the methane emission factors of cattle by studing the spatio-temporal differences in the body weight for 46 cattle species, the feeding and diet, and the draft and milk production in different places of China are also discussed . The Tier 2 equations of IPCC (2006) were used to calculate the methane emission factors from both the enteric and the manure management emissions. The calculation showed that the enteric emission factor was general low in summer (4.48 kg head-1 month-1) and high in other seasons, while the emission from manure management was high in summer (0.44 kg head-1 month-1) and low in other seasons. Spatially, northwestern China has a higher enteric methane emission factor (71.0 kg head-1 year-1) than southwestern China (55.2 kg head-1 year-1). The methane emission factor from manure management was low (0.1 kg head-1 year-1) in Tibetan plateau and high (5.4 kg head-1 year-1) in southeastern China.
Key words: cattle; methane; emission factor; spatial variation; seasonal change
BIAN Junyan , WANG Xinsheng , ZHANG Wen . An Analysis of the Spatial Variations and Seasonal Changes of Methane Emission from Cattle in China[J]. Journal of Geo-information Science, 2015 , 17(9) : 1071 -1079 . DOI: 10.3724/SP.J.1047.2015.01072
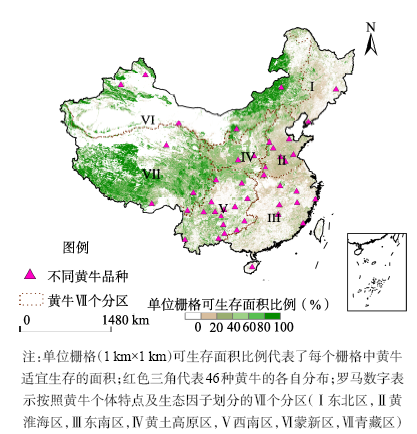
Fig. 1 Spatial distribution of different categories of cattle in China图1 黄牛亚类分布及Ⅶ个分区 |
Tab. 1 Individual characteristics of cattle in different regions of China表1 我国黄牛分区及其个体特征参数 |
| 分区 | 6个月 | 1岁 | 成年牛 | 产奶量 | 亚类 |
|---|---|---|---|---|---|
| (kg) | (kg) | (kg) | (kg/d) | ||
| Ⅰ | 122~133 | 161~232 | 425~764 | 3.8 | 延边牛、蒙古牛、复州牛 |
| Ⅱ | 76~158 | 141~255 | 310~608 | 3.0 | 渤海黑牛、冀南牛、郏县红牛、鲁西牛、蒙山牛、南阳牛、徐州牛 |
| Ⅲ | 65~133 | 106~178 | 234~601 | 3.4 | 大别山牛、广丰牛、吉安牛、锦江牛、雷琼牛、闽南牛、皖南牛、温岭高峰牛 |
| Ⅳ | 79~145 | 149~229 | 263~660 | 3.5 | 晋南牛、平陆山地牛、秦川牛、太行牛、蒙古牛 |
| Ⅴ | 41~178 | 96~211 | 214~464 | 2.0 | 巴山牛、川南山地牛、滇中牛、峨边花牛、关岭牛、黎平牛、凉山牛、南丹牛、平武牛、威宁牛、文山牛、巫陵牛、务川牛、云南高峰牛、昭通牛、隆林牛 |
| Ⅵ | 64~137 | 105~175 | 290~540 | 3.1 | 阿勒泰牛、安西牛、哈萨克牛、乌珠穆沁牛、蒙古牛 |
| Ⅶ | 50~67 | 96~110 | 190~398 | 1.6 | 柴达木牛、迪庆牛、甘孜藏牛、西藏黄牛 |
注:数据来源于中国畜禽遗传资源志·牛志[18] |
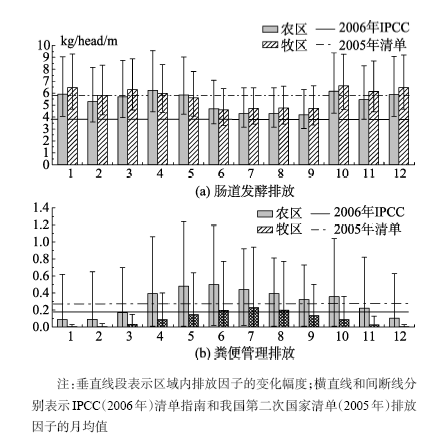
Tab. 2 Properties of cattle breeding in China and the parameters used for calculating the methane emission factors表2 我国黄牛饲养及甲烷排放相关参数季节变化 |
| 月份 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | ||
| 农区 | 规模化饲养Ym(%) | 6.5 | 6.5 | 6.5 | 6.5 | 6 | 5.5 | 5.5 | 5.5 | 5.5 | 6 | 6.5 | 6.5 |
| 散养Ym(%) | 7.0 | 7.0 | 7.0 | 7.0 | 6.5 | 6.0 | 6.0 | 6.0 | 6.0 | 6.5 | 7.0 | 7.0 | |
| DE(%) | 50 | 50 | 50 | 50 | 55 | 60 | 65 | 65 | 65 | 55 | 50 | 50 | |
| 使役时间(h) | 0.0 | 0.0 | 0.0 | 3.0 | 6.0 | 6.5 | 6.5 | 6.5 | 6.5 | 6.5 | 0.0 | 0.0 | |
| 牧区 | 规模化饲养Ym(%) | 7.0 | 7.0 | 7.0 | 7.0 | 6.0 | 5.5 | 5.5 | 5.5 | 5.5 | 6.0 | 7.0 | 7.0 |
| 散养Ym(%) | 7.5 | 7.5 | 7.5 | 7.5 | 6.5 | 6.0 | 6.0 | 6.0 | 6.0 | 6.5 | 7.5 | 7.5 | |
| DE(%) | 45 | 45 | 45 | 45 | 50 | 55 | 55 | 55 | 55 | 50 | 45 | 45 | |
| 使役时间(h) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
注:规模化标准为年出栏>50头[33] |
Fig. 2 Emission factors and the seasonal variations of the enteric and manure management systems of cattle图2 黄牛肠道发酵和粪便管理的甲烷排放因子及其季节变化 |
Fig. 3 Differences in the methane emissions from enteric and manure management systems in different regions of China图3 不同地区黄牛的肠道发酵和粪便管理甲烷排放差异 |
Tab. 3 Emission factors and the total methane emissions in each province of China表3 全国各省年甲烷排放因子(kg/head/a)及总排放量(Gg) |
| 省市 | 肠道发酵排放 | 粪便管理排放 | 省市 | 肠道发酵排放 | 粪便管理排放 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 排放因子 | 排放量 | 排放因子 | 排放量 | 排放因子 | 排放量 | 排放因子 | 排放量 | ||||
| 安徽 | 67.20 | 94.47 | 3.00 | 7.00 | 辽宁 | 65.64 | 109.70 | 1.92 | 3.00 | ||
| 北京 | 69.84 | 2.99 | 4.20 | 0.00 | 内蒙 | 69.36 | 110.69 | 3.12 | 6.00 | ||
| 福建 | 62.16 | 23.86 | 3.24 | 2.00 | 宁夏 | 67.44 | 30.49 | 0.96 | 1.00 | ||
| 甘肃 | 65.64 | 134.97 | 0.72 | 2.00 | 青海 | 63.48 | 84.58 | 0.12 | 0.00 | ||
| 广东 | 62.76 | 56.77 | 8.16 | 9.00 | 山东 | 75.24 | 375.54 | 5.64 | 39.00 | ||
| 广西 | 61.20 | 160.81 | 7.32 | 15.00 | 山西 | 69.24 | 97.74 | 3.84 | 7.00 | ||
| 贵州 | 56.64 | 192.39 | 0.72 | 3.00 | 陕西 | 71.28 | 114.06 | 0.72 | 2.00 | ||
| 海南 | 59.64 | 17.09 | 9.12 | 4.00 | 上海 | 64.20 | 0.01 | 3.12 | 0.00 | ||
| 河北 | 62.52 | 234.66 | 4.20 | 19.00 | 四川 | 60.36 | 41.29 | 0.60 | 5.00 | ||
| 河南 | 74.64 | 606.67 | 5.88 | 66.00 | 天津 | 69.36 | 10.80 | 5.16 | 0.00 | ||
| 黑龙江 | 71.16 | 150.65 | 1.56 | 3.00 | 西藏 | 57.96 | 60.80 | 0.00 | 0.00 | ||
| 湖北 | 64.08 | 61.22 | 5.76 | 10.00 | 新疆 | 75.36 | 265.21 | 0.72 | 2.00 | ||
| 湖南 | 62.28 | 113.03 | 6.12 | 17.00 | 云南 | 49.08 | 134.16 | 0.72 | 3.00 | ||
| 吉林 | 72.24 | 180.28 | 1.68 | 5.00 | 浙江 | 66.36 | 5.37 | 3.00 | 0.00 | ||
| 江苏 | 66.48 | 11.39 | 2.88 | 1.00 | 重庆 | 56.04 | 40.60 | 0.72 | 1.00 | ||
| 江西 | 61.80 | 67.91 | 6.36 | 12.00 | 总计 | 67.73 | 5638.20 | 3.24 | 244.00 | ||
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
国家畜禽遗传资源委员会.中国畜禽遗传资源志·牛志[M].北京:农业出版社,2011.
|
| [19] |
|
| [20] |
|
| [21] |
甘肃省畜牧厅,甘肃省畜禽疫病志编辑委员会.甘肃省畜禽疫病志[M].兰州:甘肃民族出版社,1992.
|
| [22] |
|
| [23] |
|
| [24] |
中国畜牧业综合区划研究组.中国畜牧业综合区划[M].北京:农业出版社,1984.
|
| [25] |
中国畜牧业年鉴编辑委员会.中国畜牧业年鉴2006 [M].北京:中国农业出版社,2006.
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}