
Journal of Geo-information Science >
Analysis of Antarctic Emperor Penguins Colonies Changes Based on Remote Sensing
Received date: 2016-11-28
Request revised date: 2017-06-20
Online published: 2017-08-20
Copyright
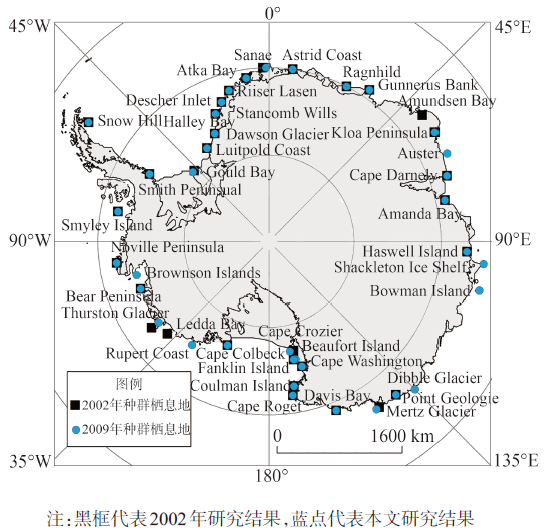
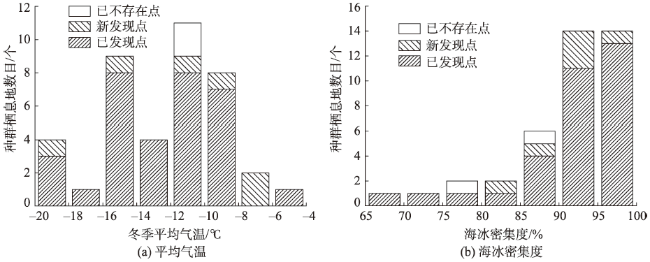
Emperor penguin is the indicator of Antarctic ecosystems. The distribution of its colonies owns essential significance for the study of Antarctic climate. Emperor penguins are sensitive to changes of the sea ice concentration and distribution. Thus, they have become an essential species for investigation on the effect of climate changes on the Antarctic ecosystems. However, it is difficult for the traditional manual investigation to obtain comprehensive and accurate information of the population colonies. Although some researchers have devoted to find emperor penguin colonies using remote sensing imageries in recent years, but their methods require considerable human involvement and cannot be used to detect all colonies rapidly and effectively. Emperor penguins breed and rest on land-fast sea ice and live in the same area for about six months, leaving extensive yellowish brown faeces which are significantly different from the white ice and snow around them. Inspired by the normalized difference snow index (NDSI), which is used for delineating snow cover, we can establish a/some similar index(s) to extract the faeces from extensive snow cover. On the basis of the difference between the reflectance of the faeces produced by emperor penguin in blue and red band, near infrared and shortwave infrared bands, two spectral indexes (NDII, EI) are putted forward to effectively recognize the faeces produced by emperor penguins, and determine their colony locations. According to the 195 scenes appropriate and quality-good Landsat 7 ETM+ imagery in 2009, a total of 38 emperor penguins colonies are obtained, 7 colonies of which are newly discovered (Bowman Island, Dibble Glacier, Auster, Point Geologie, Cape Crozier, Brownson Islands and Rupert Coast), 2 colonies have disappeared (Amundsen Bay disappeared and Ledda Bay), and the positions of the other 25 colonies (except Thuston Glacier, Luitpold, Sanae, Gould, Ragnhild and Beaufort Island) do not change significantly. The overall accuracy of colony detection is about 94% and the performance of the colony detection is influenced by the data quality and colony size. The performance of this method improves with increasing colony population size. Although spectral attributes are chosen to identify faeces produced by emperor penguins, some misclassifications may happen. This method may miss a few small colonies of which the sizes are smaller than spatial resolution of the imagery. The failure of detecting these colonies of small size is most likely due to the mixing of non-colony terrain in the Landsat pixels. These smaller colonies may well be identified by the satellites that have higher pixel resolution, and this method can be adapted to other high spatial resolution of satellite data in the future. The distribution of emperor penguin colonies is closely related to the climatic factors, and colonies tend to gather at the regions where temperature is low and ice concentration is high. It is different for climate change occurring at different colonies. Long-time and regional observations are needed to study the relationship between climate and the changes to the distribution of colonies. With the continued rise of the air temperature and the change of ice concentration, the colonies of which the latitude are below 70 °S are facing greater threat, and emperor penguin population shows a trend to shrink to the southern pole.
Key words: Emperor penguin; population colonies; change; detecton; climate; Antarctica
SHEN Xiaoyi , KE Changqing , ZHANG Jie . Analysis of Antarctic Emperor Penguins Colonies Changes Based on Remote Sensing[J]. Journal of Geo-information Science, 2017 , 19(8) : 1132 -1140 . DOI: 10.3724/SP.J.1047.2017.01132
Fig. 1 Flowchart of this method图1 方法流程图 |
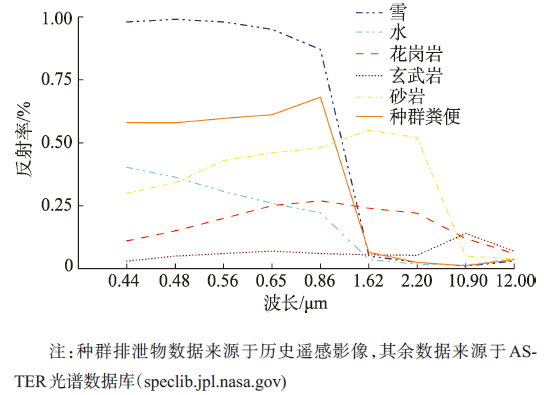
Fig. 2 Spectral reflectance of major ground objects in Antarctica图2 南极主要地物光谱反射率 |
Fig. 3 One example about areas covered by faeces in real-color image and the extraction result图3 帝企鹅种群排泄物真彩色影像和提取结果示例 |
Tab. 1 Details information of emperor penguin colonies in 2009表1 2009年南极主要帝企鹅种群栖息地 |
| 地点 | 经度 | 纬度 | 影像时间(月/年) | 与2002年位置的距离/km |
|---|---|---|---|---|
| Cape Colbeck | 158°W | 77°S | 10/2009 | 1.49 |
| Rupert Coast | 143°W | 75°S | 10/2008 | |
| Thuston Glacier | 126°W | 74°S | 10/2009 | 10.32 |
| Bear Peninsula | 110°W | 74°S | 11/2009 | 3.27 |
| Brownson Islands | 104°W | 74°S | 11/2009 | |
| Noville Peninsula | 98°W | 72°S | 11/2009 | 0.00 |
| Smyley | 79°W | 72°S | 11/2009 | 1.16 |
| Smith | 61°W | 74°S | 10/2009 | 1.26 |
| Snowhill | 57°W | 65°S | 10/2009 | 0.96 |
| Gould | 48°W | 78°S | 10/2009 | 9.14 |
| Luitpold | 34°W | 77°S | 11/2009 | 21.50 |
| Dawson | 27°W | 76°S | 10/2009 | 3.16 |
| Halley | 27°W | 76°S | 10/2009 | 6.77 |
| Stancomb | 23°W | 74°S | 10/2009 | 4.93 |
| Drescher | 19°W | 73°S | 10/2009 | 7.95 |
| Riiser | 15°W | 72°S | 10/2009 | 2.33 |
| Atka | 8°W | 71°S | 09/2009 | 1.11 |
| Sanae | 1°W | 70°S | 10/2009 | 49.72 |
| Princess Astrid Coast | 8°E | 70°S | 11/2009 | 1.12 |
| Ragnhild | 27°E | 70°S | 10/2009 | 8.62 |
| Gunnerus | 34°E | 69°S | 10/2009 | 3.59 |
| Kloa Point | 57°E | 67°S | 10/2009 | 0.68 |
| Auster | 64°E | 67°S | 10/2009 | |
| Cape Darnley | 70°E | 68°S | 10/2009 | 0.16 |
| Amanda Bay | 77°E | 69°S | 10/2009 | 0.12 |
| Haswell Island | 93°E | 67°S | 08/2009 | 1.11 |
| Shackleton Ice Shelf | 96°E | 65°S | 10/2009 | |
| Bowman Island | 103°E | 65°S | 10/2009 | |
| Dibble Glacier | 135°E | 66°S | 10/2009 | |
| Point Geologie | 140°E | 67°S | 10/2009 | 0.44 |
| Mertz Glacier Tongue | 147°E | 67°S | 11/2009 | 8.48 |
| Davis Bay | 158°E | 69°S | 10/2009 | 3.86 |
| Cape Washington | 165°E | 75°S | 10/2009 | 1.15 |
| Beaufort Island | 167°E | 76°S | 10/2009 | 60.07 |
| Franklin Island | 168°E | 76°S | 10/2009 | 0.80 |
| Cape Crozier | 169°E | 77°S | 10/2009 | |
| Coulman Island | 170°E | 73°S | 10/2009 | 1.44 |
| Cape Roget | 171°E | 72°S | 10/2009 | 1.55 |
Fig. 4 Spatial distribution of 38 emperor penguin colonies in 2009 in this study图4 2009年南极38个帝企鹅种群栖息地的空间分布 |
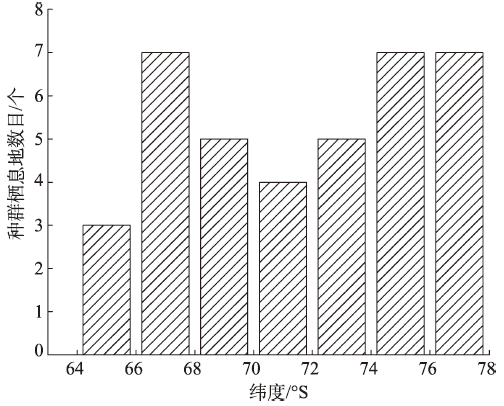
Fig. 5 Distribution of emperor penguin colonies along latitude in 2009图5 2009年南极帝企鹅种群纬度位置的分布 |
Fig. 6 Distribution of average air temperature and ice concentration in emperor penguin colonies in winter图6 南极帝企鹅种群位置的冬季平均气温和冬季海冰密集度分布 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
[
|
| [9] |
[
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
[ Zhao Z Y, Liu Z, Gong P. Automatic extraction of floating ice at Antractic continental margin from remotely sensed imagery using object-based segmentation[J]. Sci China Earch Sci, 2012,1:8. ]
|
| [22] |
[
|
| [23] |
|
| [24] |
Hardisky,
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
[
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}