
Journal of Geo-information Science >
Assessment of Polar Bear Habitats Stability from Remote Sensing
Received date: 2018-01-16
Request revised date: 2018-06-13
Online published: 2018-09-25
Supported by
National Key Research and Development Program of China, No.2018YFC1407203.
Copyright
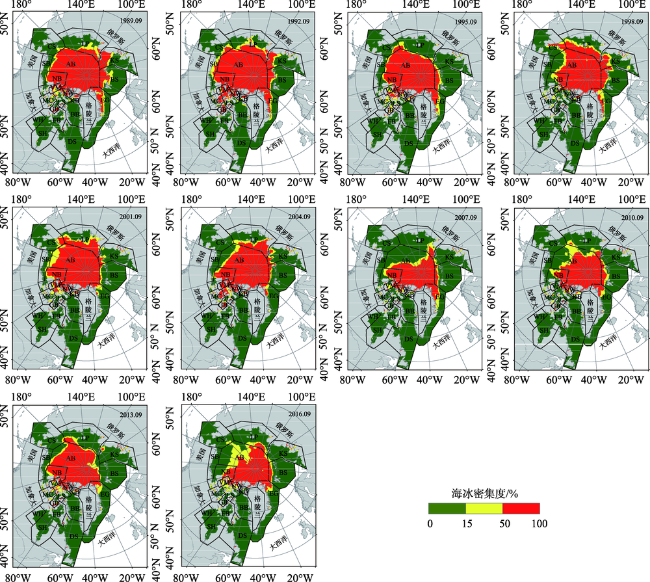
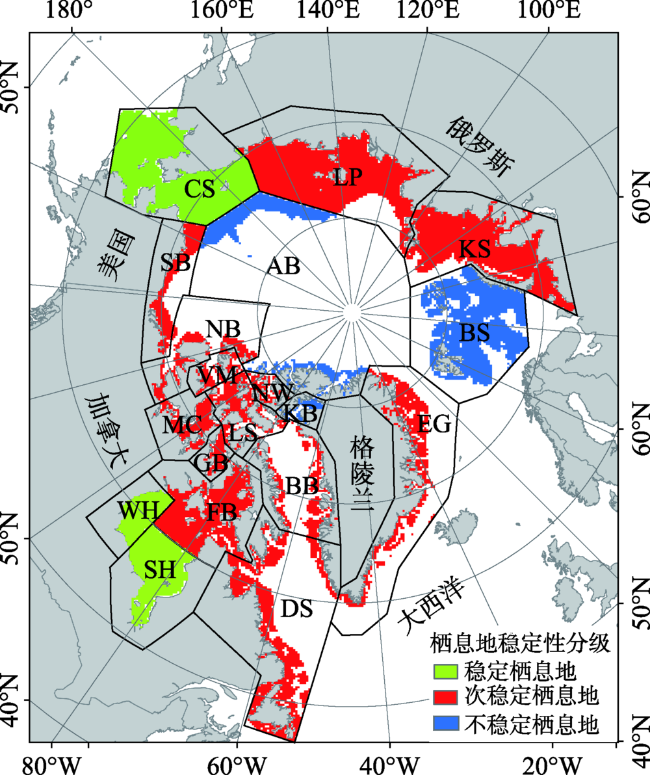
Polar bear is one of the most important mammals in the Arctic, but its number decreased in recent years. Polar bears are sensitive to changes of the sea ice distribution and depend on sea ice as a platform for hunting, moving and reproducing. In other words, sea ice is an important part of polar bear habitat. Climate is the main factor of sea ice changes. Therefore, it is very important to understand the current situation of polar bears as well as the effect of climate on the Arctic ecosystem. Although many researchers have devoted to find polar bears habitat using aerial survey in recent years, their methods require considerable human involvement and cannot be used to detect all habitats rapidly and effectively. Thus, it is necessary to find a method to quickly assess the polar bear habitat changes. Based on the sea ice concentration products from the United States National Snow and Ice Data Center (NSIDC) and the ETOPO1 bedrock product provided by the NOAA, the inter-annual variability of sea ice concentration, open water area, sea ice retreat, sea ice advance and the length of the open water season in the Arctic were analyzed. Then, the polar bear habitat stability were analized. The results indicate that from 1989 to 2016 the sea ice concentration has decreased, open water area increased and multiyear ice decreased. Most of the multiyear ice has converted to one-year ice. The sea ice appeared later and retreated earlier, so the length of the open water season increased significantly, an increase of 72 days compared to 1992. Barents Sea is the region with the most significant changes in open water area and the length of open water season among 19 habitats, with increasing rates of 9.71×103 km2/a and 71.69 days/decade, respectively. Based on the change rates of the length of the open water season, we divide the polar bears habitats into three levels of conditions: stable, sub-stable and instable. The three stable habitats, including the Chukchi sea, Western Hudson Bay and Southern Hudson Bay are located in the lower latitudes compared with other habitats. There are 13 sub-stable habitats, including Laptev Sea, Kara Sea, East Greenland, Baffin Bay, Davis Strait, Foxe Basin, Gulf of Boothia, M’Clintock Channel, Viscount Melville, Norwegian Bay, Northern Beaufort Southern Beaufort and Lancaster Sound. The three unstable habitats are located in the north of 70°N, including Arctic Basin, Barents Sea and Kane Basin. Stable habitats are mainly in low latitudes, and unstable regions are all in high latitudes. The classification results show that the high latitude area is covered with more sea ice, but the inter-annual variation is very significant. In three unstable regions, polar bears have less time to adapt to the sea ice changes, and the inter-annual migration changes greatly, which is less favorable to the survival and development of polar bears.
Key words: Polar bear; open water; habitat; stability; Arctic
LI Haili , KE Changqing . Assessment of Polar Bear Habitats Stability from Remote Sensing[J]. Journal of Geo-information Science, 2018 , 20(9) : 1327 -1337 . DOI: 10.12082/dqxxkx.2018.180057
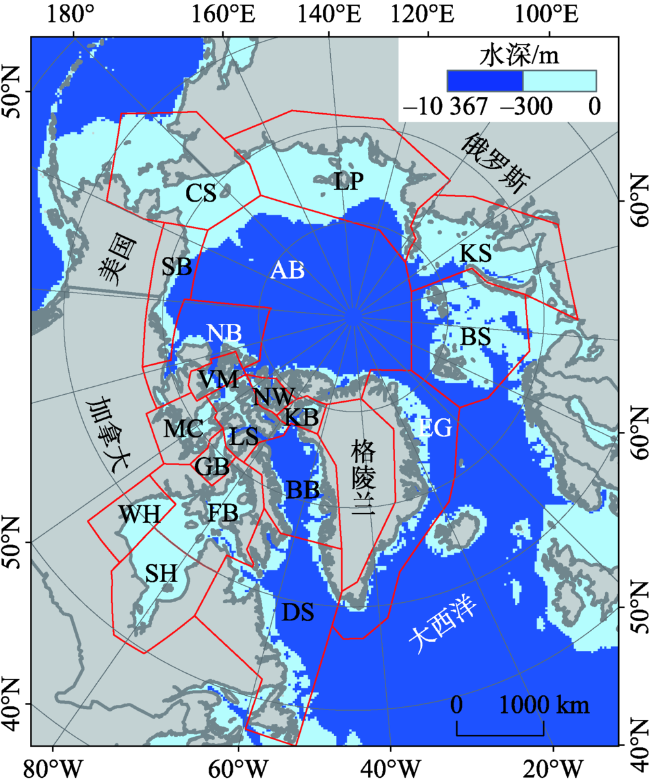
Fig.1 Location and water depth of the 19 polar bear habitats图1 19个北极熊栖息地及水深分布 |
Tab.1 Habitats in English and Chinese表1 栖息地中英文对照 |
| 序号 | 英文全称 | 英文简写 | 中文名称 | 序号 | 英文全称 | 英文简写 | 中文名称 |
|---|---|---|---|---|---|---|---|
| 1 | Kane Basin | KB | 凯恩盆地 | 11 | Western Hudson Bay | WH | 西哈得孙湾 |
| 2 | Baffin Bay | BB | 巴芬湾 | 12 | Southern Hudson Bay | SH | 南哈得孙湾 |
| 3 | Lancaster Sound | LS | 兰开斯特海峡 | 13 | Davis Strait | DS | 戴维斯海峡 |
| 4 | Norwegian Bay | NW | 挪威湾 | 14 | East Greenland | EG | 东格陵兰 |
| 5 | Viscount Melville | VM | 梅尔维尔子爵海峡 | 15 | Barents Sea | BS | 巴伦支海 |
| 6 | Northern Beaufort | NB | 北波弗特 | 16 | Kara Sea | KS | 喀拉海 |
| 7 | Southern Beaufort | SB | 南波弗特 | 17 | Laptev Sea | LP | 拉普捷夫海 |
| 8 | M’Clintock Channel | MC | 麦克林托克海峡 | 18 | Chukchi Sea | CS | 楚科奇海 |
| 9 | Gulf of Boothia | GB | 布西亚湾 | 19 | Arctic Basin | AB | 北极盆地 |
| 10 | Foxe Basin | FB | 福克斯湾 |
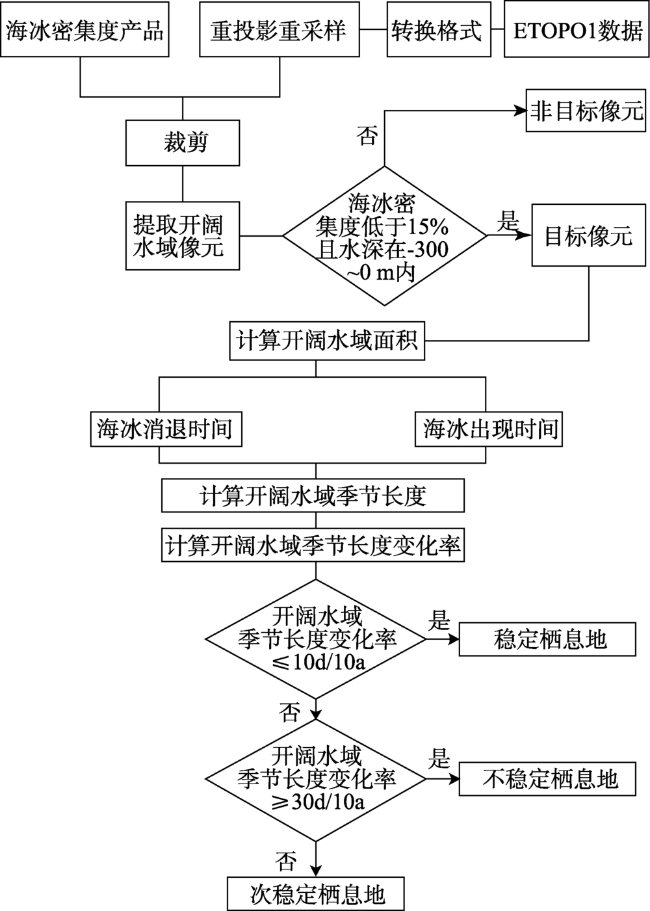
Fig. 2 Flowchart for determining the stability of the polar bear habitat图2 北极熊栖息地稳定性判断流程图 |
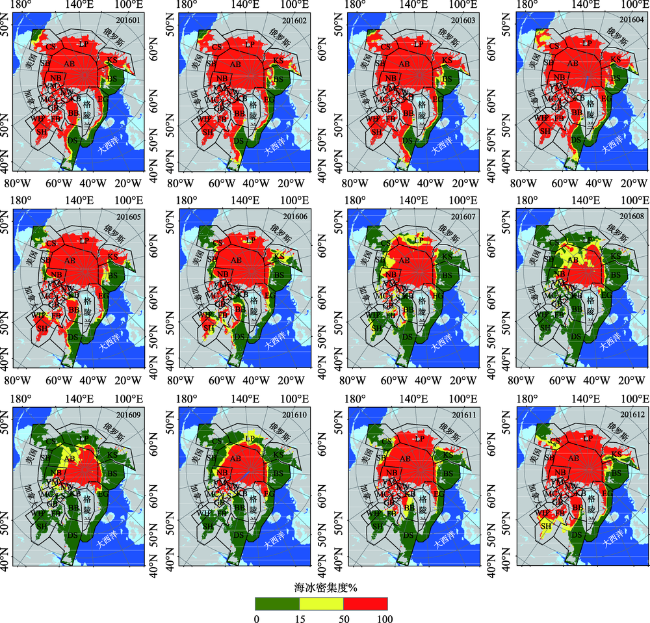
Fig. 3 Monthly variation of sea ice concentration in 2016图3 2016年海冰密集度月变化 |
Fig. 4 Sea ice concentration in September from 1989 to 2016图4 1989-2016年9月海冰密集度变化 |
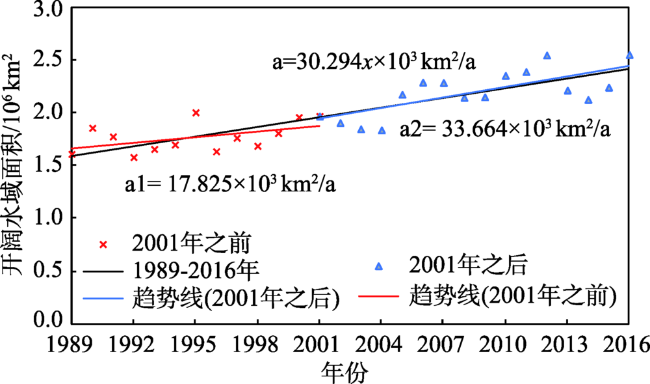
Fig. 5 The annual change of open water area and subparagraph of its slope in North polar region图5 北极开阔水域面积变化及斜率分段 |
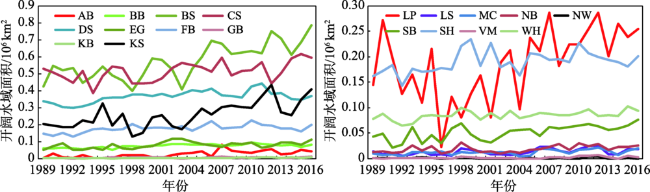
Fig. 6 The annual change of open water area in the 19 polar bear habitats图6 19个北极熊栖息地开阔水域面积变化 |
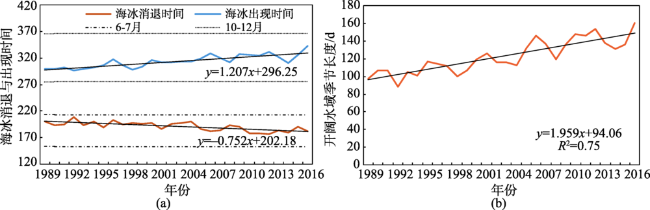
Fig. 7 The day of the year (DOY) of the initial sea ice retreat and advance and the length of the open water season in the North Polar region图7 北极海冰消退、出现时间和开阔水域季节长度 |
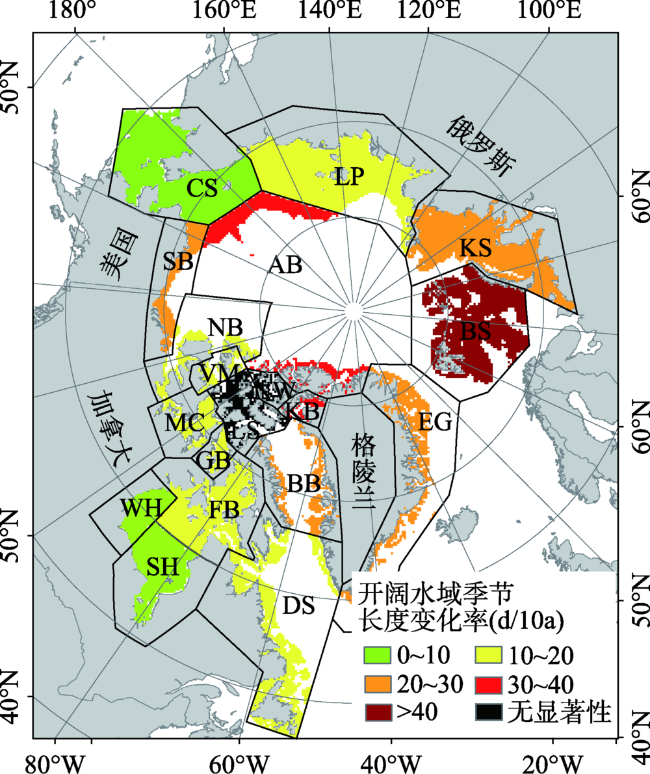
Fig. 8 Changing rate of the length of the open water season for polar bear habitat图8 北极熊栖息地开阔水域季节长度变化率 |
Tab. 2 The length of the open water season and its change rate in 19 polar bear habitats表2 19个北极熊栖息地开阔水域季节长度及变化率 |
| 栖息地 | 均值/d | 标准差/d | 变化率/(d/10a) | 可决系数R2 |
|---|---|---|---|---|
| AB | 48.25 | 35.84 | 31.88 | 0.54** |
| BB | 163.93 | 32.23 | 24.94 | 0.41** |
| BS | 174.79 | 79.52 | 71.69 | 0.55** |
| CS | 176.39 | 16.42 | 9.81 | 0.24** |
| DS | 202.00 | 23.46 | 14.83 | 0.27** |
| EG | 84.14 | 34.56 | 24.74 | 0.35** |
| FB | 135.68 | 17.50 | 11.54 | 0.27** |
| GB | 40.50 | 25.03 | 13.51 | 0.20* |
| KB | 33.75 | 36.88 | 33.84 | 0.57** |
| KS | 104.36 | 29.91 | 24.54 | 0.46** |
| LP | 70.57 | 25.01 | 14.04 | 0.21* |
| LS | 35.54 | 25.55 | 9.66 | 0.10 |
| MC | 50.21 | 24.30 | 12.76 | 0.19* |
| NB | 95.71 | 26.82 | 14.00 | 0.18* |
| NW | 13.29 | 19.61 | 0.74 | 0.00 |
| SB | 94.18 | 28.64 | 22.31 | 0.41** |
| SH | 152.32 | 16.37 | 7.72 | 0.15* |
| VM | 22.00 | 23.57 | 14.64 | 0.26** |
| WH | 149.43 | 15.96 | 9.38 | 0.23** |
注:*在0.05水平上显著相关;**在0.01水平上显著相关 |
Fig.9 Stability assessment result for the habitats图9 北极熊栖息地稳定性评估结果 |
The authors have declared that no competing interests exist.
| [1] |
[
|
| [2] |
|
| [3] |
|
| [4] |
[
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
[
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
[
|
| [23] |
|
| [24] |
[
|
| [25] |
|
| [26] |
[
|
| [27] |
[
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
[
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}