
Journal of Geo-information Science >
Study on the Relationship between Terrain and Distribution of the Vegetation in Shennongjia Forestry District
Received date: 2019-09-27
Request revised date: 2019-11-14
Online published: 2020-05-18
Supported by
National Science and Technology Basic Resource Investigation Program(2017FY100900)
Copyright
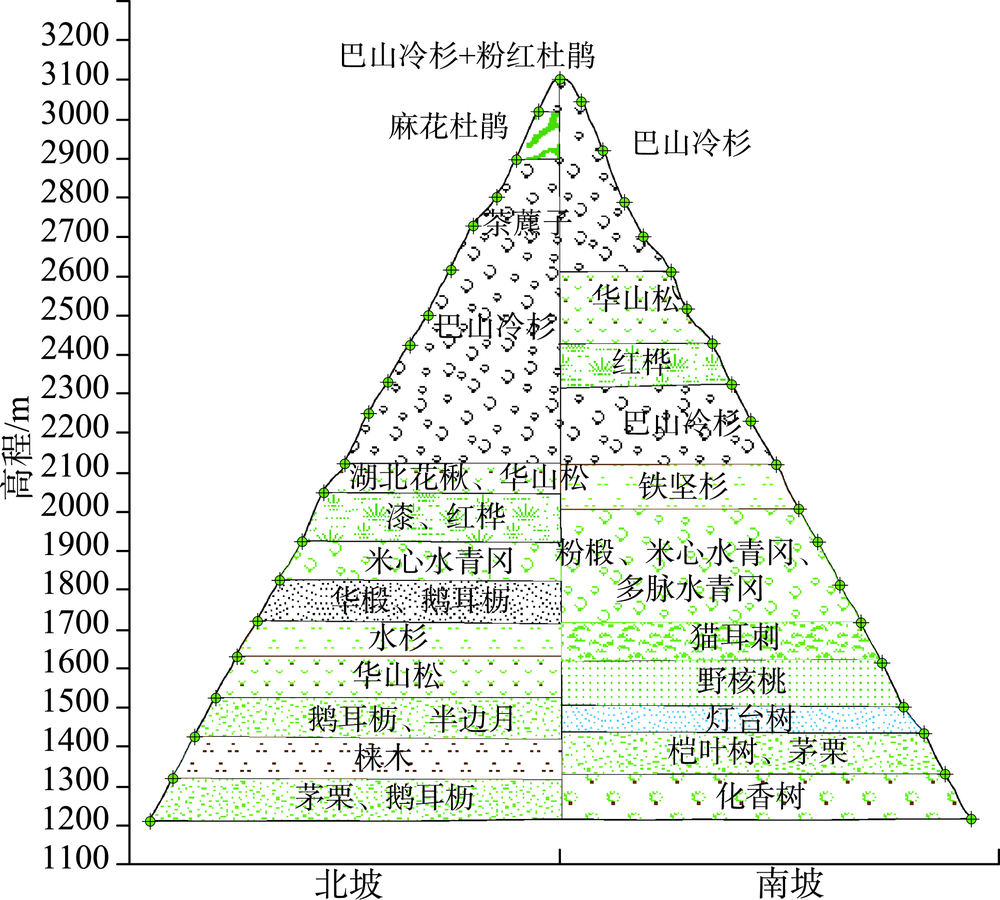
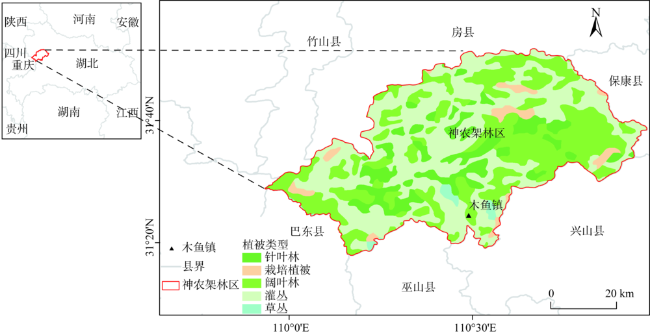
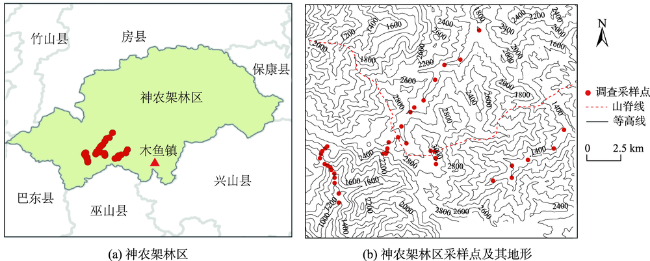
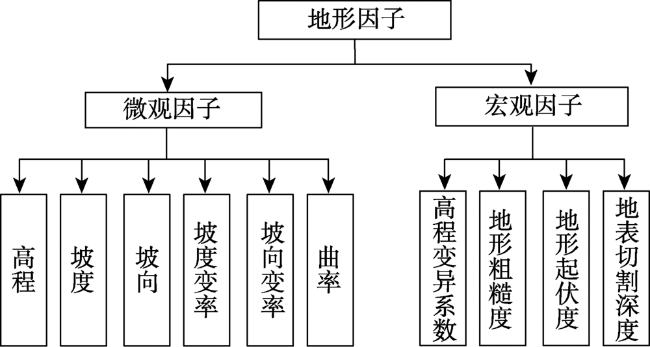
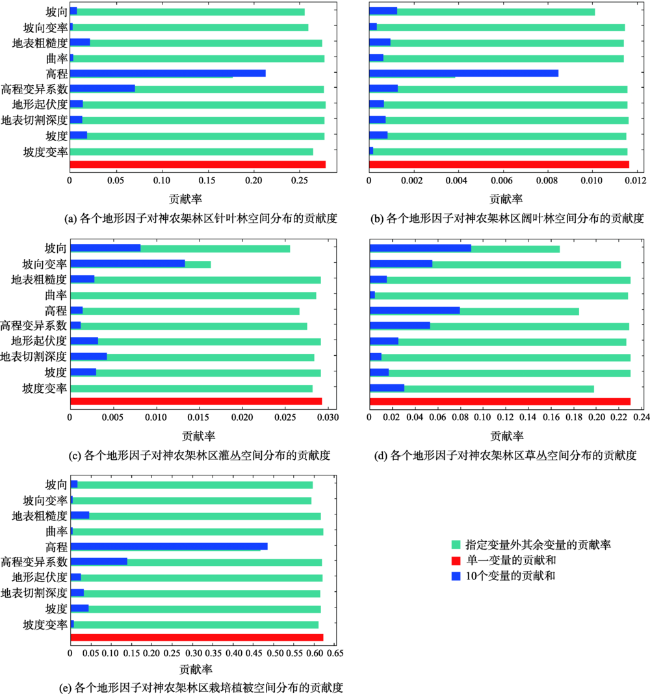
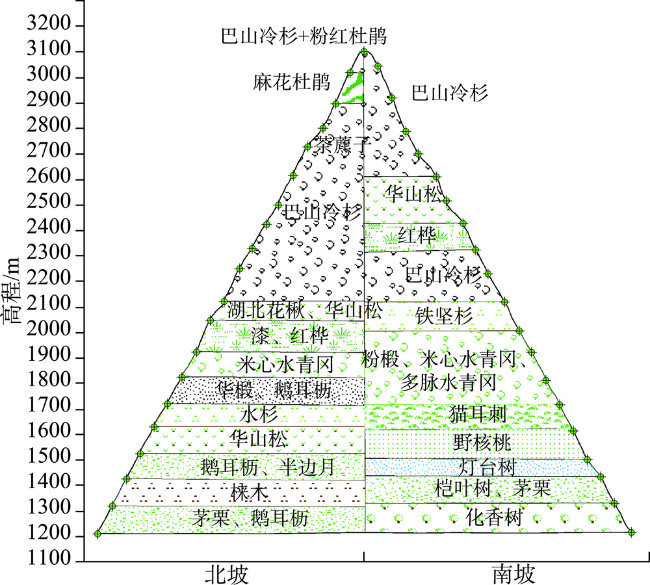
The Shennongjia Forestry District is one of the areas with the highest biodiversity in China. The complex topography exerts great influence on vegetation distribution in this region. This paper used the maximum entropy model (MaxEnt), digital elevation data, vegetation distribution map, and field-surveyed data, to study how the topographic characteristics affect local typical vegetation distributions at two scales, i.e., vegetation type and population levels. The relationship models between vegetation type and topographic factors, and between plant population and topographic factors were established respectively by quantifying the topographic ranges of vegetation types and plant species. Results show that: (1) the spatial distribution of different vegetation types was affected by different topographic factors. The distribution of coniferous forests was affected by elevation and coefficient of variation in elevation, the distribution of broad-leaved forests was controlled by elevation and aspect, and the distribution of shrubs was controlled by aspect and the slope of aspect. The factors affecting the distribution of grasses were various. (2) The elevation ranges of typical plant species were generally consistent with those of vegetation types. Specifically, 90% of coniferous forests were distributed at elevation between 1600 and 2600 m, and the typical populations of Abies fargesii and Pinus armandii were distributed at elevation of 1700~3200 m. 85% of the broad-leaved forests were distributed within the range of 1000 m to 2000 m in elevation, and the typical populations of Cyclobalanopsis glauca, Carpinus turczaninowii concentrated at elevation between1000 to 2000 m. 95% of shrubs occurred at slope of aspect within 0~40 degrees, and the typical populations of Rhododendron simsii and Rosaceae mainly occurred at slope of aspect less than 40 degrees. The relationship models used for vegetation types and plant species were different. The relationship between vegetation types and topographic factors was fitted using Gauss model. While the relationship between typical species and topographic factors was relatively complex, and the distribution patterns of different species were even different. (3) Vegetation distribution showed a rather weak relationship with typical slope characteristic. This study provides a basic reference for vegetation protection, vegetation restoration, and vegetation management in the Shennongjia region.
ZHANG Xinghang , ZHANG Baiping , WANG Jing , YAO Yonghui , YU Fuqin . Study on the Relationship between Terrain and Distribution of the Vegetation in Shennongjia Forestry District[J]. Journal of Geo-information Science, 2020 , 22(3) : 482 -493 . DOI: 10.12082/dqxxkx.2020.190553
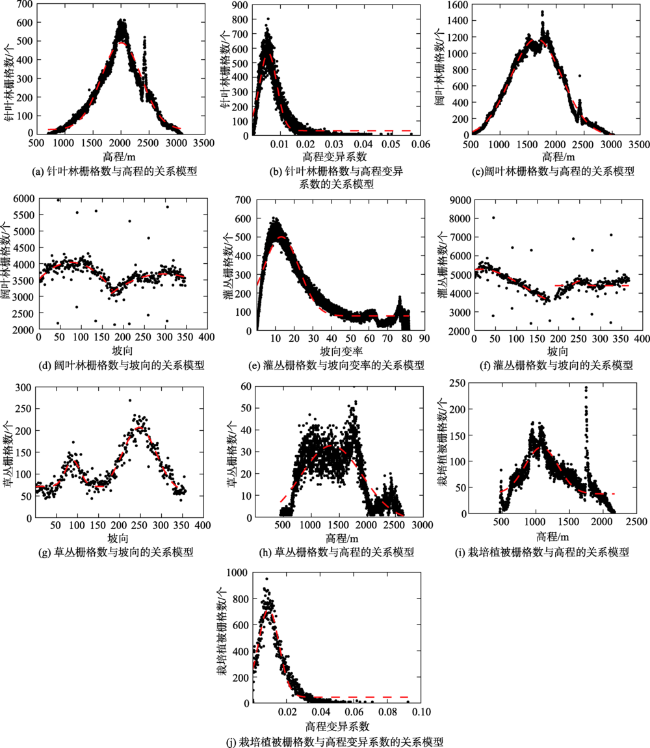
表1 神农架林区植被类型与地形因子关系模型的拟合参数Tab. 1 Fitting parameters of relationship model between vegetation types and topographic factors in Shennongjia Forestry District |
| 关系模型 | 拟合参数 | ||||
|---|---|---|---|---|---|
| a | b | c | w | /% | |
| 针叶林与高程 | 464.5 | 2005.7 | 25.3 | 356.2 | 94.2 |
| 针叶林与高程变异系数 | 537.3 | 0.0056 | 32.4 | 0.003 | 92.8 |
| 阔叶林与高程 | 1189.9 | 1646.0 | -14.3 | 457.6 | 97.9 |
| 阔叶林与坡向(第一个峰) | 3 054 761.1 | 76.0 | -3 050 725.7 | 4171.6 | 33.2 |
| 阔叶林与坡向(第二个峰) | 989 602.2 | 304.5 | -985 918.5 | 3557.6 | 22.1 |
| 灌丛与坡向变率 | 423.9 | 13.1 | 77.5 | 9.5 | 63.1 |
| 灌丛与坡向(第一个峰) | 2301.6 | 14.8 | 2980.4 | 100.5 | 62.6 |
| 灌丛与坡向(第二个峰) | — | — | — | — | — |
| 草丛与坡向(第一个峰) | 59.8 | 88.7 | 71.3 | 17.6 | 66.9 |
| 草丛与坡向(第二个峰) | 148.5 | 246.9 | 58.4 | 40.2 | 89.1 |
| 草丛与高程 | 34.4 | 1361.3 | -1.6 | 545.7 | 65.2 |
| 栽培植被与高程 | 89.3 | 1085.8 | 37.6 | 244.4 | 62.4 |
| 栽培植被与高程变异系数 | 672.0 | 0.0093 | 45.7 | 0.006 | 93.8 |
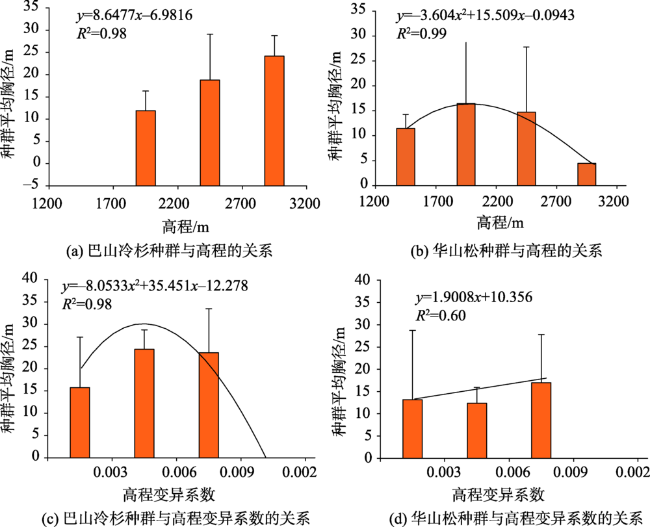
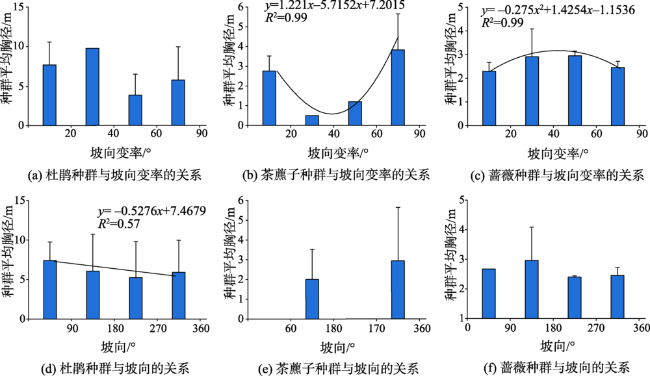
图7 神农架调查样地针叶林典型植被种群与高程和高程变异系数的关系Fig. 7 Relationship between typical vegetation population and altitude and altitude variability coefficient of coniferous forest in Shennongjia |
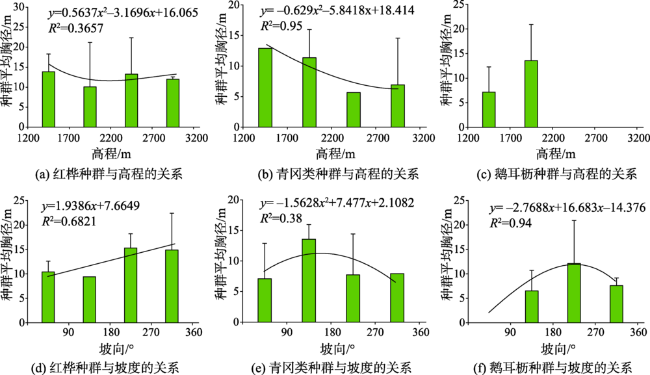
图8 神农架调查样地阔叶林典型植被种群与高程和坡向的关系Fig. 8 Relationship between typical vegetation populations and elevation and slope direction of broad-leaved forest in Shennongjia |
| [1] |
伍光和, 蔡运龙 . 综合自然地理学.第2版[M]. 北京: 高等教育出版社, 2004.
[
|
| [2] |
黄秉维 . 现代自然地理[M]. 北京: 科学出版社, 2004.
[
|
| [3] |
程东亚, 李旭东 . 喀斯特地区植被覆盖度变化及地形与人口效应研究[J]. 地球信息科学学报, 2019,21(8):1227-1239.
[
|
| [4] |
汤国安, 李发源, 刘学军 . 数字高程模型教程[M]. 北京: 科学出版社, 2010.
[
|
| [5] |
|
| [6] |
薛凯凯, 熊礼阳, 祝士杰 , 等. 基于DEM的黄土崾岘提取及其地形特征分析[J]. 地球信息科学学报, 2018,20(12):1710-1720.
[
|
| [7] |
宋朝枢 . 神农架自然保护区科学考察集[M]. 北京: 中国林业出版社, 1999.
[
|
| [8] |
沈茂才 . 中国秦岭生物多样性的研究和保护[M]. 北京: 科学出版社, 2010.
[
|
| [9] |
廖明尧 . 神农架地区自然资源综合调查报告[M]. 北京: 中国林业出版社, 2015.
[
|
| [10] |
马旭东, 张苏峻, 苏志尧 , 等. 车八岭山地常绿阔叶林群落结构特征与微地形条件的关系[J]. 生态学报, 2010,30(19):5151-5160.
[
|
| [11] |
张容, 董廷发, 邓晓保 , 等. 西双版纳20公顷样地热带森林植被数量分类与排序[J]. 生态学杂志, 2018,37(2):347-352.
[
|
| [12] |
张梅玲, 徐睿择 . 定南县岭北矿区植被覆盖与地形因子关系分析[J]. 地理信息世界, 2018,25(4):50-55.
[
|
| [13] |
赵鹏, 屈建军, 徐先英 , 等. 长江源区沙化高寒草地植被群落特征及其与地形因子的关系[J]. 生态学报, 2019,39(3):1030-1040.
[
|
| [14] |
李亚飞, 刘高焕 . 大香格里拉地区植被空间分布的环境特征[J]. 自然资源学报, 2011,26(8):1353-1363.
[
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
张金屯 . 数量生态学[M]. 北京: 科学出版社, 2004.
[
|
| [21] |
|
| [22] |
|
| [23] |
张东方, 张琴, 郭杰 , 等. 基于MaxEnt模型的当归全球生态适宜区和生态特征研究[J]. 生态学报, 2017,37(15):5111-5120.
[
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
丁军军 . 神农架森林土壤微生物沿海拔分布格局及形成机制[D]. 北京:清华大学, 2016.
[
|
| [29] |
中国科学院《中国植被图》编辑委员会. 中华人民共和国植被图(1:100万)[M]. 北京: 地质出版社, 2007.
[ Editorial committee of vegetation map of China, CAS. Vegetation map of the People's Republic of China (1:1 000 000) [M]. Beijing: Geological Publishing House, 2007. ]
|
| [30] |
刘爱利 . 基于1:100万DEM的我国地形地貌特征研究[D]. 西安:西北大学, 2004.
[
|
| [31] |
井学辉, 曹磊, 郭仲军 , 等. 阿尔泰山小东沟林区植被随地形分布规律[J]. 应用与环境生物学报, 2015,21(3):152-158.
[
|
| [32] |
|
| [33] |
|
| [34] |
杨帆 . 基于DEM形态分析的湖南丘陵地形景观策略研究[D]. 长沙:湖南大学, 2015.
[
|
| [35] |
|
| [36] |
|
| [37] |
王毅, 郭跃 . 喀斯特地貌区植被覆盖与地形因子的空间关系分析——以贵州普定县为例[J]. 长江流域资源与环境, 2018,27(1):157-167.
[
|
| [38] |
胡俊忠, 张镱锂, 于海彬 . 基于MaxEnt模型和GIS的青藏高原紫花针茅分布格局模拟[J]. 应用生态学报, 2015,26(2):505-511.
[
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}